The plumulaceous (downy) and pennaceous feathers of Pennaraptora/Paraves evolved from Pterosaur pycnofibres and actinofibrils.
Pycnofibres covered the body, while the actinofibrils were covered by the wing membrane.Pycnofibres are comparable to Stage II feathers that have unfurled out of their sheaths.
Actinofibrils are comparable to Stage IIIa feathers that are still within their sheaths.
Actinofibrils
http://en.wikipedia.org/wiki/Pterosaur
While historically thought of as simple, leathery structures composed of skin, research has since shown that the wing membranes of pterosaurs were highly complex and dynamic structures suited to an active style of flight. The outer wings (from the tip to the elbow) were strengthened by closely spaced fibers called actinofibrils.[17] The actinofibrils themselves consisted of three distinct layers in the wing, forming a crisscross pattern when superimposed on one another.
The variation of space between adjacent actinofibrils in Jeholopterus, also reported in Rhamphorhynchus (Padian & Rayner 1993), suggests that those fibres were connected by some elastic tissue that enabled them to spread apart or join whenever necessary, making the actinopatagium more flexible (perhaps somewhat elastichttps://archosaurmusings.wordpress.com/2009/08/05/what-on-earth-are-pycnofibers/
For the first time we observe actinofibrils (those fibers that support the wings) lying in multiple layers (not just a single one) and these tend to cross each other a little, though they are essentially subparallel. This tells us something about the structure and to a lesser extent function of the wing.
https://archive.org/stream/The_Pterosaurs_From_Deep_Time_by_David_M._Unwin/The_Pterosaurs_From_Deep_Time_by_David_M._Unwin_djvu.txt
In this [Sordes], as in other pterosaurs, the wing fibers were embedded within the patagia [wing membranes] and typically measured a little less than one-tenth of a millimetre in diameter- about twice the thickness of a human hair. In some spots unravelled fibers reveal that they were composite structures composed of at least 20 or 30 very fine strands, wound together in a helical fashion. Each strand was only a few hundredth of a millimeter across and probably made of collagen a material that is common in the skin of vertebrates".http://www.ucmp.berkeley.edu/vertebrates/flight/pter.html
Bird feathers are analogous to the wing fibers of pterosaurshttp://www.springerlink.com/content/v8434087565413kk/fulltext.pdf?page=1
Wellnhofer [4, 5] and Padian [6, 7], following von Zittel [8],described a system of fine structural fibers investing the wing membrane, in a pattern similar to the orientation of the feather shafts of birds and the wing fingers of bats, both principal structural elements supporting the patagium and responsible for the transmission of aerodynamic force.
http://www.jstor.org/pss/2400656
The wing membrane was supported and controlled through a system of stiffened, intercalated fibers,which were oriented like the main structural elements in the wings of birds and bats.https://archosaurmusings.wordpress.com/2008/06/16/pterosaur-wings-2-structure/
Actinofibrils are unusual structures and we are not sure exactly what they are composed of. The best guess is collagen, but it could also be cartilage or keratin. Determining this in fossils is obviously near impossible but all three are realistic possibilities, though of course collagen is the most likely given the position of the fibres inside the wing membrane (rather than on the surface) and they do not connect to the bones of the wing finger. They lie sub-parallel to the wing towards the wingtips and then sub-perpendicular as we move more proximally. There are no actinofibrils in the proximal wing close to the body, and they get more densely packed the further away you go.https://www.google.ca/url?sa=t&rct=j&q=&esrc=s&source=web&cd=2&cad=rja&uact=8&ved=0ahUKEwjs7aOI4OzQAhUI42MKHd4DCfAQFgggMAE&url=https%3A%2F%2Fwww.fhsu.edu%2Facademic%2Fresearch%2Freports%2FBennettSabbaticalAbstractF12%2F&usg=AFQjCNH5up6EVndVPejQhmiCw_TJqRhuOQ&sig2=clu3jNdQyuxYHSZTPoXd9Q&bvm=bv.141320020,d.cGc
Actinofibrils were formed in place by incremental addition of layer upon layer of hard keratin.
http://prumlab.yale.edu/sites/default/files/prum_n_brush_2002.pdf
Stage II — Origin of a collar with differentiated barb ridges results in a mature feather with a tuft of unbranched barbs and a basal calamus emerging from a superficial sheath.
Stage IIIa — Origin of helical displacement of barb ridges and the new barb locus results in a pinnate feather with an indeterminate number of unbranched barbs fused to a central rachis.

https://en.wikipedia.org/wiki/Scansoriopteryx#Feathers_and_scales
One distinctive feature of Scansoriopteryx is its elongated third finger, which is the longest on the hand, nearly twice as long as the second finger (in most theropod dinosaurs, the second finger is the longest). This is unlike the configuration seen in most other theropods, where the second finger is longest. The long wing feathers, or remiges, appear to attach to this long digit instead of the middle digit as in birds and other maniraptorans. Shorter feathers are preserved attached to the second finger.[6] A relative of Scansoriopteryx, Yi, suggests that this elongated third finger supported a membranous wing of some kind alongside feathers.[7]
Pycnofibres
http://www.dinosaur-museum.org/featheredinosaurs/rhamphorhynchoid.pdf
A NEW RHAMPHORHYNCHOID WITH A HEADCREST AND COMPLEX INTEGUMENTARY STRUCTURES
STEPHEN A. CZERKAS and QIANG JI
The [rhamphorhynchoid] “hair-like” structures [pycnofibres]
are also unique in being preserved in fully three
dimensionally forms as compared to two
dimensional staining or impressions. The hairs
[pycnofibres] are shown to be complex multi-strand structures instead of single strands or actual hairs. The complex nature
of these filaments most closely resembles natal
down feathers, but apparently without having
barbules. As such, they may represent the earliest
known form of feathers. This implies that such
integumentary structures may have originated
independently among pterosaurs from that of birds,
or that birds and pterosaurs may share a common
ancestor which had evolved this kind of insulation
before fight had been achieved in either group.
Feathers differ significantly from hair in that their multiple strands, the barbs,emanate from a single hollow structure, called the calamus. The integumentary structures seen in Pterorhynchus [a rhamphorhynchid] bear a striking similarity to that of a natal down feather with only the notable absence of having the additional barbules branching from the barbs. This absence is significant all the more because without the barbules, the barbs emanating from a calamus represents the hypothetical “Stage II” structure speculated as being an incipient step in the evolution of feathers (Prum, 1999).Proto-feathers have been attributed to two pterosaurs which are of similar animals (Ji and Yuan, 2002; Wang, et al., 2002). Even more so, the morphology details seen in Pterorhynchus demonstrate that the integumentary structures of pterosaurs are not like hair, but are analogous to being proto-feathers. Specifically, they resemble natal down feathers where individual filaments are seen to spread from a single follicle.
Therefore, the individual filaments are not representative of hair,but are analogous to being the barbs of a feather.Barbules, if present, cannot be discerned whichsuggests that they either did not exist, or that thelimits of preservation have obscured them.Nonetheless, the morphology of having several
barbs stemming from a short calamus indicates that
the body covering of Pterorhynchus are feather homologues. Without barbules, these structures would represent the second stage of feather
development as speculated by Prum (1999). Thefeather homologues of Pterorhynchus alsodemonstrate that a primary function achieved bythese plumulaceous feathers was that of thermalinsulation, and that feathers with a true rachis andbarbs aligned into well developed vanes representa derived condition.
The wing membranes are thought to have been stiffened by internal fibers, called aktinofibrils (Martill and Unwin, 1989; Wellnhofer, 1987, 1991). The distal end of a wing membrane is preserved in Pterorhychus which shows clear aktinofibrils that are aligned in parallel rows.http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2842671/
Kellner et al
On the tenopatagium close to the body and on the tail, a third type of fibre [pycnofibre] with somewhat diffuse edges is observed (figures 3a and and44a). Type C fibres can be easily separated from other fibres by their dark-brown colour and their general lack of organization. They are distributed along the body, the tail and the tip of the actinopatagium close to the fourth wing finger phalanx (figures 1, ,22 and and44c). Sometimes clustering together, they are not found covering the external portion of the plagiopatagium and are apparently rare on the actinopatagium.
As Wang et al. (2002) pointed out, these fibres are best interpreted as structures covering the body, commonly referred to as ‘hair’ or hair-like structures (e.g. Sharov 1971; Bakhurina & Unwin 1995). This pterosaur hair, which is not homologous to the mammalian hair (a protein filament that originates deep in the dermis and grows through the epidermis), is here called pycnofibre (from the Greek word pyknos, meaning dense, bushy). The pycnofibres are further formed by smaller fibrils of unknown nature. They were possibly mostly composed of keratin-like scales, feathers and mammalian hair.
Two other Chinese specimens were reported with integumental covering, coming from the same stratum (the Daohugou Bed) as Jeholopterus. So far we have not had the opportunity to examine this material. The first one is a small unnamed anurognathid with extensive preservation of soft tissue, including fibres that have been interpreted as protofeathers (Ji & Yuan 2002). The published pictures show that the soft tissue interpreted as protofeathers is of the same nature as the pycnofibres of Jeholopterus. There is no indication of branching structures that are expected for feather precursors.
http://pterosaurnet.blogspot.ca/2014/01/helpful-background.html
http://pterosaurnet.blogspot.ca/2011/10/bird-feathers-are-analogous-to-wing.html
http://pterosaurnet.blogspot.ca/2010/05/pterosaur-wing-covering-2.htmlThe strings cross the fibres at angles between 30° and 90°
https://pterosaurheresies.wordpress.com/2016/03/page/2/
Several views of the Pterorhynchus wingtip. The ungual (manual4.5) has been colored magenta. Manual 4.4 (green) is broken.

https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2842671/
Although no distinctive trailing edge is discernible, the wing membrane extends along the body and is connected to the hind limbs, reaching the ankle (Wang et al. 2002). While the distal portion of the plagiopatagium shows several layers of closely packed fibres (actinofibrils), the more proximal part lacks these structures. This confirms the observations of Schaller (1985), who recognized two distinct portions of the plagiopatagium, the actinopatagium and the tenopatagium, distinguished by the presence and absence of actinofibrils, respectively.
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4202321/
Compared with contour feathers, flight feathers have a larger pennaceous vane and a longer and thicker rachis. Wing flight feathers also have a longer calamus for insertion deeper into the follicle and anchor more securely to sustain its aerodynamic function
http://www.peteducation.com/article.cfm?c=15+1829&aid=2776
The bases of the flight feathers are covered with smaller contour feathers called coverts. There are several layers of coverts on the wing.
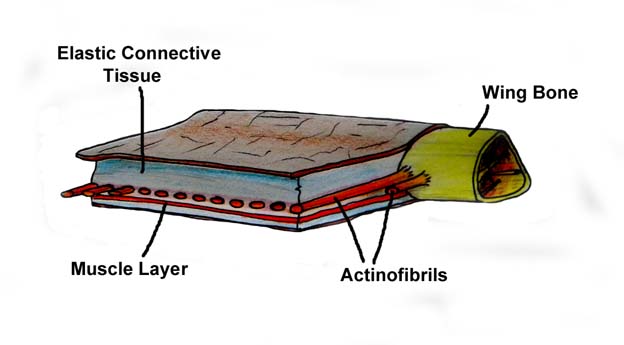
Both pycnofibres and actinofibrils are the same basic form. But they differ in that the pycnofibres proceeded through the stages to where the sheaths had disintegrated and the internal strands are visible on the surface. The actinofibrils did not proceed to that point. In the actinofibril, the internal strands (barbs) are still within the sheaths. And the sheaths are covered by the wing membrane.
The pycnofibres are on the body of the pterosaur. When the sheath disintegrates they unfurled into a form comparable to natal down and kept the body of the pterosaur warm.
The actinofibrils perform a different function. They stiffen and strengthen the wing membrane.
It is helpful to keep in mind that the actinofibrils and the membrane grew in synch as the pterosaur grew. (It is an example of facilitated variation.)
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4386666/
In a fractal-like fashion, other morphogenetic processes take place within the barb plate to form numerous barbules (Figure 2c; Table 1).
https://royalsocietypublishing.org/cms/attachment/c4afbb49-1ec8-4e5a-b40d-891d96dfea8c/rspb20091980f05.jpg

A schematic view of the three major structural components of the feather rachis. (a) (i) superficial layers of *fibres, the ultimate size-class in the hierarchy of feather keratin filaments (approx. 6 µm diameter), wound circumferentially round the rachis. (ii) The majority of the fibres extending parallel to the rachidial axis and through the depth of the cortex. Part of the section is peeled back to show why the fibres and even megafibrils are not usually recognized in histological sectioning, but rather only fibrils lower down the hierarchy (based on the electronic supplementary material, figure S2c). Any longitudinal section along the line of the arrows or at any point along the height of the fibre other than at the fibre surface (arrowheads) will fail to show the fibre. (iii) It shows the medulloid pith comprising gas-filled polyhedral structures (based on SEM images, electronic supplementary material, figures S5 and S6). Inset, part of a steel rebar with nodes, used in engineering technology to reinforce high-rise structures, analogous to rachidial fibres. (b) Schematic cross section of fibres and biodegraded ‘matrix’: (i) fibres; (ii) residual cytosol of keratinocytes presumably housing effete organelles and perhaps cytoskeletal elements—all degraded along with corneous envelope; (iii) interdigitating plasma membrane of the original keratinocytes with associated corneous envelope proteins. (c) A schematic three-dimensional cross section of the rachis showing approximate thickness (based on SEMs) of the three keratin layers comprising, (i) circumferential and (ii) longitudinal fibres of the cortex and (iii) polyhedra of medulloid pith. Asterisk denotes homologous with syncitial barbules.
http://prumlab.yale.edu/sites/default/files/prum_n_brush_2002.pdf
Figure 2. Schematic Diagram of Helical Growth of Barb Ridges of a Pennaceous Feather The branched structure of the barbs and the rachis of a feather form by helical growth and fusion of barb ridges within the tubular feather germ. Feathers grow from the base. Barb ridges form at the new barb locus on the posterior midline of the collar and grow helically around the collar toward the anterior midline where they fuse to form the rachis ridge. Subsequent barb ridges fuse to the rachis ridge. In feathers with an afterfeather, the new barb locus divides into two laterally displaced new barb loci. Subsequently, new barb ridges grow helically both anteriorly to the main rachis and posteriorly to form the hyporachis and vane of the afterfeather. The main vane and the afterfeather form separate vanes united within a single feather by the calamus (Figure 1A). Pennaceous feathers obtain their planar form only after emerging from the cylindrical feather sheath when growth is complete. The obverse (upper) and reverse (lower) surfaces of the vane develop from the outer and inner surfaces of the cylindrical feather germ. Illustration based on Lucas and Stettenheim (1972).
https://www.youtube.com/watch?v=_8ykumFZXFQ
Down Feather Development: Gallus domestics
Dinosaur Museum
http://dinosaur-museum.org/featheredinosaurs/arboreal_maniraptoran.pdf
AN ARBOREAL MANIRAPTORAN FROM NORTHEAST CHINA STEPHEN A. CZERKAS and CHONGXI YUAN
http://www.dinosaur-museum.org/featheredinosaurs/rhamphorhynchoid.pdf
A NEW RHAMPHORHYNCHOID WITH A HEADCREST AND COMPLEX INTEGUMENTARY STRUCTURES
STEPHEN A. CZERKAS and QIANG JI
AND
http://www.springer.com/gp/about-springer/media/springer-select/dinosaurs-are-not-the-great-great-grandparents-of-birds-/29304
https://waxing-paleontological.blogspot.ca/2015/05/scansoriopterygid-faq.html?showComment=1482173666510#c1515808862453816302
Feduccia and Czerkas
http://www.scielo.cl/scielo.php?script=sci_arttext&pid=S0717-95022007000100010
https://blogs.scientificamerican.com/tetrapod-zoology/yi-qi-is-neat-but-might-not-have-been-the-black-screaming-dino-dragon-of-death/




http://veteriankey.com/avian-deflighting-techniques/

https://books.google.ca/books?id=livLBAAAQBAJ&pg=PA18&lpg=PA18&dq=humeral+patagium&source=bl&ots=COa0_l7raO&sig=XSQ8a63WrZ1qHXhZg-g12C0rito&hl=en&sa=X&ved=0ahUKEwiY95S3mo3RAhUiwYMKHfZLAgoQ6AEIIzAC#v=onepage&q=humeral%20patagium&f=false
https://books.google.ca/books?id=KG86AgWwFEUC&pg=PA128&lpg=PA128&dq=bird+propatagium+feathers+rooted+in+tendon&source=bl&ots=RuJ5ZnWI1K&sig=B_bTFKkAMp5pM73VxpgyWdEtncg&hl=en&sa=X&ved=0ahUKEwib6tfiu5LRAhUM7YMKHYsQAwEQ6AEIGzAA#v=onepage&q=bird%20propatagium%20feathers%20rooted%20in%20tendon&f=false (Page 128)
An alternate way to understand the pterosaur wing:
The pterosaur postpatagium helps to hold and support the actinofibrils in place.
http://rspb.royalsocietypublishing.org/content/277/1679/321.full
https://www.youtube.com/watch?v=_8ykumFZXFQ
Down Feather Development: Gallus domestics
Dinosaur Museum
http://dinosaur-museum.org/featheredinosaurs/arboreal_maniraptoran.pdf
AN ARBOREAL MANIRAPTORAN FROM NORTHEAST CHINA STEPHEN A. CZERKAS and CHONGXI YUAN
http://www.dinosaur-museum.org/featheredinosaurs/rhamphorhynchoid.pdf
A NEW RHAMPHORHYNCHOID WITH A HEADCREST AND COMPLEX INTEGUMENTARY STRUCTURES
STEPHEN A. CZERKAS and QIANG JI
AND
http://www.springer.com/gp/about-springer/media/springer-select/dinosaurs-are-not-the-great-great-grandparents-of-birds-/29304
https://waxing-paleontological.blogspot.ca/2015/05/scansoriopterygid-faq.html?showComment=1482173666510#c1515808862453816302
See how it's covered in skin already? There's a tendon running between the shoulder and the wrist, just like in pterosaurs, that anchors a skin membrane called the propatagium. The ulna is covered in thick skin that anchors the flight feathers. In many birds, the bottom of the upper arm is loosely connected to the body by skin as well. So you can imagine these membranes becoming larger and more parachute-like.
Czerkas & Yuan also noticed tissue impressions coming off the ulna and third finger of Scansoriopteryx—those might be better interpreted (now) as a flight membrane.https://www.researchgate.net/publication/271658034_Jurassic_archosaur_is_a_non-dinosaurian_bird
Feduccia and Czerkas
There are indications from where the feathers emanate below the ulna which suggest that a short patagium may have been present. Unlike most of the wing feathers, there appears to be a series of feathers that do not reach the bone itself.
http://www.scielo.cl/scielo.php?script=sci_arttext&pid=S0717-95022007000100010
In barb medullary cells feather keratin is accumulated in few peripheral bundles that merge with those of cortical cells to form the wall of the ramus. The latter is joined with branching barbules.
https://blogs.scientificamerican.com/tetrapod-zoology/yi-qi-is-neat-but-might-not-have-been-the-black-screaming-dino-dragon-of-death/
What’s been mostly overlooked in discussions of Yi qi is that pennaraptoran maniraptorans already have patagia.
Look at the (perhaps familiar) pictures of nightjar wings below and observe all the ‘webbing’ that surrounds the fingers and arm. A membrane called the propatagium spans the space between the wrist and shoulder, and membranes run along the trailing (or posterior, or postaxial) edges of the hand and ulna too.
Xu et al. (2015) also note that these [Yi qi] patches have an unusual wrinkled texture, not typical of skin that would have been covered in filaments or feathers. But maybe they’re naked and wrinkled for taphonomic reasons.
As a Tet Zoo regular, you'll recognise these images from p. 19 of Katrina van Grouw's The Unfeathered Bird. They show the left wing of a European nightjar (Caprimulgus europaeus) in (at top) dorsal and ventral views. Note all the skin membranes around the more muscular parts of the wing. Image by Katrina van Grouw, used with permission.
http://veteriankey.com/avian-deflighting-techniques/
The four patagia of the wing include the propatagium, where the wing and the neck join the thorax; the postpatagium, which is located at the caudal angle of the carpus; the metapatagium at the caudal junction of the thorax and the wing, and the alular patagium between the alula and the carpometacarpus.
https://books.google.ca/books?id=livLBAAAQBAJ&pg=PA18&lpg=PA18&dq=humeral+patagium&source=bl&ots=COa0_l7raO&sig=XSQ8a63WrZ1qHXhZg-g12C0rito&hl=en&sa=X&ved=0ahUKEwiY95S3mo3RAhUiwYMKHfZLAgoQ6AEIIzAC#v=onepage&q=humeral%20patagium&f=false
"humeral patagium"
https://books.google.ca/books?id=KG86AgWwFEUC&pg=PA128&lpg=PA128&dq=bird+propatagium+feathers+rooted+in+tendon&source=bl&ots=RuJ5ZnWI1K&sig=B_bTFKkAMp5pM73VxpgyWdEtncg&hl=en&sa=X&ved=0ahUKEwib6tfiu5LRAhUM7YMKHYsQAwEQ6AEIGzAA#v=onepage&q=bird%20propatagium%20feathers%20rooted%20in%20tendon&f=false (Page 128)
propatagium and postpatagiumhttps://en.wikipedia.org/wiki/Flight_feather
Remiges (from the Latin for "oarsman") are located on the posterior side of the wing. Ligaments attach the long calami (quills) firmly to the wing bones, and a thick, strong band of tendinous tissue known as the postpatagium helps to hold and support the remiges in place.[2]
An alternate way to understand the pterosaur wing:
The pterosaur postpatagium helps to hold and support the actinofibrils in place.
http://rspb.royalsocietypublishing.org/content/277/1679/321.full
On the tenopatagium close to the body and on the tail, a third type of fibre with somewhat diffuse edges is observed (figures 3a and 4a). Type C fibres can be easily separated from other fibres by their dark-brown colour and their general lack of organization. They are distributed along the body, the tail and the tip of the actinopatagium close to the fourth wing finger phalanx (figures 1, 2 and 4c). Sometimes clustering together, they are not found covering the external portion of the plagiopatagium and are apparently rare on the actinopatagium.http://prumlab.yale.edu/sites/default/files/prum_1999_mde_development.pdf
Generally thicker than the actinofibrils (figure 4a), type C fibres have an average thickness ranging between 0.2 and 0.5 mm. In several places, it is clear that they are formed by smaller fibrils, the nature of which is unknown. The sediment between type C fibres tends to be light brown in colour, making the distinction of individual fibres more difficult. Several cross each other but lack the reticular pattern formed by the multi-layered superposition of the actinofibrils. In several areas, type C fibres are preserved associated with an amorphous whitish matter that has been interpreted as patches of the epidermis (and dermis, described earlier). Although not parallel to each other and lacking the organization of the actinofibrils, fibres C in most parts are generally displaced away from the skeleton.
As Wang et al. (2002) pointed out, these fibres are best interpreted as structures covering the body, commonly referred to as ‘hair’ or hair-like structures (e.g. Sharov 1971; Bakhurina & Unwin 1995). This pterosaur hair, which is not homologous to the mammalian hair (a protein filament that originates deep in the dermis and grows through the epidermis), is here called pycnofibre (from the Greek word pyknos, meaning dense, bushy). The pycnofibres are further formed by smaller fibrils of unknown nature. They were possibly mostly composed of keratin, like scales, feathers and mammalian hair.
One detail, however, of feather development appears to violate von Baer’s rule. During the development of the first feather papillae in the embryo (before day 12 in the chick, Gallus gallus), the barb ridge primordia appear as longitudinal condensations within the feather papillae before the follicle and collar are fully formed (Lucas and Stettenheim, ’72). However, this developmental event—the origin of the feather before the follicle and collar—is clearly derived because barb ridges would be unable to grow without the spatial organization provided by the collar.
ReplyDeletehttps://archosaurmusings.wordpress.com/2009/08/05/what-on-earth-are-pycnofibers/
David Hone
03/11/2010 at 3:19 pm
Yes, that’s a typo. It should of course read more as “….keratin, like scales, feather and hair”.
https://bio.unc.edu/files/2011/04/FeducciaCzerkas2015.pdf
ReplyDeleteWe focus here on the discovery of an anatomical feature,
the propatagium, which argues that Caudipteryx
supports the neoflightless hypothesis—that is, it is derived
from a flighted ancestry (Paul 2002)—and therefore its
highly derived avian anatomy was selected for in an
aerodynamic context.
http://prumlab.yale.edu/sites/default/files/prum_1999_mde_development.pdf
ReplyDeleteAvian bristles are characterized by the increased
strength of the rachis and the reduction (sometimes
complete) of the barbs and barbules. Bristles
evolved through the derived reduction in barb
number and barbule structure within the follicle
(stage Vf).
hooklet = barbicel = hamuli
ReplyDeletehttps://archosaurmusings.wordpress.com/2008/06/16/pterosaur-wings-2-structure/
ReplyDeleteActinofibrils are unusual structures and we are not sure exactly what they are composed of. The best guess is collagen, but it could also be cartilage or keratin. Determining this in fossils is obviously near impossible but all three are realistic possibilities, though of course collagen is the most likely given the position of the fibres inside the wing membrane (rather than on the surface) and they do not connect to the bones of the wing finger. They lie sub-parallel to the wing towards the wingtips and then sub-perpendicular as we move more proximally. There are no actinofibrils in the proximal wing close to the body, and they get more densely packed the further away you go.
https://archosaurmusings.wordpress.com/2009/08/05/what-on-earth-are-pycnofibers/
ReplyDeleteHowever being, well, pterosaurs one could hardly call their ‘fur’ fur, or hair for that matter, these being the preserve of synapsids (and by extensions, mammals). Nor (despite some calls for it) could they be considered protofeathers as there is no established homology between them and saurischian protofeathers and feathers (or for that matter ornithischian dermal structures). This has left pterosaur researchers with a problem – these are clearly different things and we refer to the quite often but with no-one having gone out onto a limb and named them, we were left with a variety of half-names all of them convoluted or presented in quotation marks.
http://www.theodora.com/encyclopedia/f/feather.html
ReplyDeletec, Two barbules of the posterior series - seen only in crosssection in b. The rods comprising the lateral fringe, or vane, are known as the rami or the " barbs," and will be found, on microscopic examination, to be lath-shaped and to taper to a point. Further, each barb supports a double series of smaller outgrowths known as the radii, or "barbules "; so that each barb may be likened to a feather in miniature. These " barbules," however, differ markedly in structure on the two sides of the barb, those pointing towards the tip of the feather - the " anterior barbules " - being ribbon-shaped from the base outwards for about half their length, when they become cut up to form a series of long and very delicate hooklets (fig. I'd). On the opposite side of the barb the barbules are also ribbon-shaped for about half their length, but the ribbon is curved trough-fashion, so that the whole series of posterior barbules forms a number of deep valleys, and into these the 'hooklets are thrust so as to catch hold of the upper edges of the troughs, which are set so that the f, Feather-Barbs.
https://academy.allaboutbirds.org/feathers-article/2/
ReplyDeleteThe wing feathers specialized for flight are characterized by uniform windproof surfaces, or vanes, on either side of the central shaft that are created by an interlocking microstructure. Also called remiges
, these feathers are asymmetric with a shorter, less flexible leading edge that prevents midair twisting.
http://ncsce.org/pages/feathers.html
ReplyDeleteAlthough a feather resembles a tree (with a trunk sprouting branches and smaller branches sprouting from those branches), the way a feather grows is quite different from the way a tree grows. Feather growth is more akin to the story of the way a sculptor produces a figure from a block of marble - the statue of David was already in the rock, Michelangelo simply removed anything that wasn't David. Similarly, a feather does not start as a shaft which sprouts branches, it begins as a tapered cylinder and cell-death creates the structures.
ReplyDeleteA very important perception of the pterosaur wing membrane is that it is a set of fibres (actinofibrils) which are joined together by a skin membrane.
https://pterosaur.net/restoration.php
ReplyDeleteMany fossils also preserve hair-like integument, which is often called "ptero-fuzz". Ptero-fuzz varied from coarse to fine, and seems to have been extensive in at least some pterosaurs. The fuzz may have been dry and truly "fuzzy" as in bats, or it may have been kept oily and slicked, as bird feathers. Certainly have an oily coat would have helped prevent fishing pterosaurs from getting water-logged if they came in contact with the water.
Similarities:
ReplyDeleteActinofibril and feather rachis
Patagium
Multiple layers and overlapping feathers
http://www.nature.com/articles/srep14864
ReplyDeleteParticularly remarkable is the [Enantiornithes] presence of an almost completely preserved postpatagium, extending from the caudoproximal base of the ungual phalanx of digit II to the preserved proximal-most caudal portion of the ulna and in clear association with the wing’s plumage (Fig. 1). The preserved calami of flight feathers (i.e., primaries and secondaries) are embedded in the postpatagium, forming the same sinuous caudal outline visible in the naked (i.e., unfeathered) wing of modern birds10,17
https://pterosaur.net/ecology.php
ReplyDeleteThe fibrolamellar texture of pterosaur bone suggests that they had high metabolic rates akin to those of birds and mammals and were therefore capable of rapid growth, but size distributions clusters of single pterosaur species (notably those of the Jurassic form Rhamphorhynchus) suggest their growth rate was low: Rhamphorhynchus may have taken several years to achieve its full size (Fig. 1). This is quite unlike the growth rates of the metabolically supercharged birds and mammals, so what gives?
I am glad you like it. Care to contribute?
ReplyDelete